Papel da succinilacetona como fonte endógena de radicais livres e espécies tripletes
Leandro O. Royer (PG), Fernanda S. Knudsen (PG) e Etelvino J.H. Bechara (PQ)
Departamento de Bioquímica, Instituto de Química, Universidade de São Paulo
Palavras-chave: espécies tripletes, radicais livres, succinilacetona.
INTRODUÇÃO:
A succinilacetona (SA) é um catabólito de Tyr que se acumula nas tirosinemias, doenças caracterizadas por danos renais, hepáticos e neurológicos. Além disso, leva ao acúmulo do ácido 5-aminolevulínico (ALA), um potente agente pró-oxidante em porfirias , devido à forte ação inibitória da SA sobre 5-aminolevulínico desidratase (ALA-D). A SA também está implicada na alta freqüência de hepatomas devido à sua ação inibitória sobre a DNA ligase. Ratos tratados com ALA ou SA sofrem descarga de ferro e estresse oxidativo.
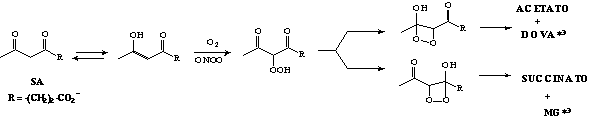
A SA é uma b-dicetona cuja forma enólica é passível de oxidação unieletrônica por oxidantes tais como peroxinitrito (ONOO-) e hemeproteínas. O radical enoil formado pode sofrer inserção de oxigênio resultando em dioxetano intermediário, cuja clivagem produz espécies carbonílicas tripletes. O longo tempo de vida da espécie triplete (ms) a capacita participar de processos bimoleculares, reagindo ou transferindo energia de excitação eletrônica. Estas espécies, se geradas a nível celular, podem desempenhar papéis úteis ou deletérios à célula. Além dos radicais e espécies tripletes gerados, a oxidação aeróbica de SA tem como produtos principais o ácido 4,5-dioxovalérico (DOVA) e o metilglioxal (MG), aldeídos reconhecidamente citotóxicos e genotóxicos (Esquema 1).
Esquema 1
OBJETIVOS:
Investigar uma possível via mecanística de lesão hepática ou neurológica em porfirias e tirosinemias por intermediários reativos (radicais e espécies tripletes) gerados pela oxidação aeróbica de succinilacetona.
MÉTODOS:
O consumo de oxigênio por SA, iniciada por peroxinitrito ou citocromo c foi seguido em oxígrado. As reações iniciadas por peroxinitrito tiveram a velocidade de reação bem como o efeito de pH estudados espectrofotometricamente pela técnica de “stopped-flow”. Os estudos de quimiluminescência (CL) foram realizados em um luminômetro. A detecção de radicais livres intermediários foi feita por EPR utilizando MNP como captador. Metilglioxal e DOVA foram detectados por HPLC através de sua derivatização com 1,2-diaminobenzeno.
RESULTADOS:
A oxidação da SA (0,5 mM) iniciada por peroxinitrito (0,5 mM) em tampão fosfato pH 7.2 consome todo o oxigênio do meio em alguns segundos. Esse consumo responde a variações nas concentrações de SA e ONOO- bem como à concentração de tampão. Esse último dado indica o enol como substrato da oxidação uma vez que fosfato catalisa a enolização de compostos carbonílicos. O perfil de pH obtido através da oximetria e “stopped-flow” apontam o ânion peroxinitrito como o oxidante. A intensidade da emissão aumenta com [ONOO-], o mesmo não ocorrendo com a [SA]. A intensidade maxima de CL começa a cair com o aumento da concentração de SA. A presença do sensibilizador [Ru(bpy)3]2+ não só aumenta a intensidade da emissão como também a torna proporcional à concentração de SA. O espectro de EPR obtido apresenta um sexteto, indicando a presença de um radical secundário intermediário como esperado. Metilglioxal e DOVA foram detectados como produtos principais. Utilizando-se o sistema citocromo c/H2O2 como oxidante, obteve-se um consumo total de oxigênio muito semelhante ao obtido com peroxinitrito, embora a cinética seja mais lenta. A ausência de CL e a presença de um sinal triplete no espectro de EPR sugerem outro mecanismo de reação.
CONCLUSÕES:
O consumo de oxigênio, CL direta e indireta, radicais detectados por EPR e análise de produtos evidenciou o possível envolvimento de dioxetanos na oxidação aeróbica desta b-dicetona. Estes intermediários, uma vez gerados, podem levar a formação de espécies tripletes, cuja presença foi sugerida pela CL associada ao consumo de oxigênio. Estes intermediários podem ser responsáveis por alguns dos efeitos tóxicos da deficiência metabólica da tirosina.
BIBLIOGRAFIA:
1.-Lindblad, B., Lindstedt, S., & Steen, G. (1977) Proc. Natl. Acad. Sci USA 74, 4641-4645; 2.-Prieto-Alamo, M.J. & Laval, F. (1998) Proc. Natl. Acad. Sci. USA 95, 12614-12618; 3.-Bechara, E.J.H. (1996) Braz. J. Biol. Med. Res. 29, 841-851; 4.-Rocha, M.E.M., Bandy, B., Costa, C.A., Barros, M.P., Pinto, A.M.P., & Bechara, E.J.H. (1999) Free Rad. Res., no prelo; 5.-Cilento, G. (1995) Ci. Cult. 47, 312-319.
FAPESP